Auteurs :
Anne Sophie Armand, Enseignante-Chercheuse en biologie, Université Paris Cité
Raphaël Baye, Ostéopathe
Christophe Chiquet, Ostéopathe
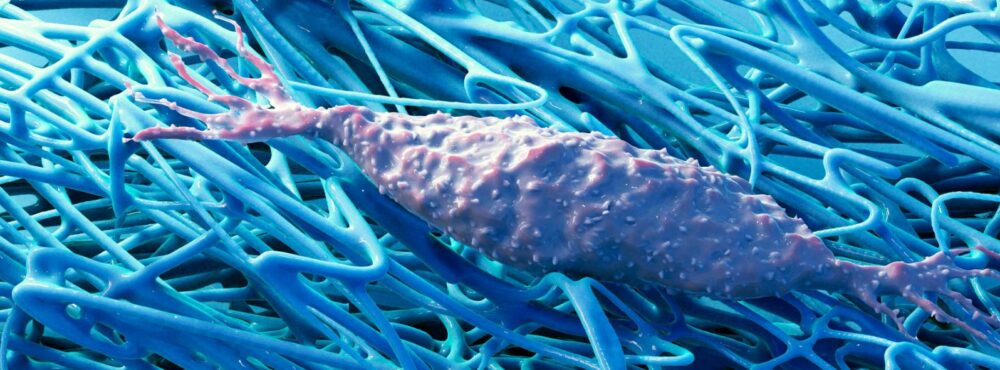
Les thérapies manuelles, largement utilisées en rééducation ou en traitement de première instance comme l’ostéopathie, la chiropractie ou la manipulation fasciale, ont longtemps cherché à établir leur légitimité scientifique en faisant appel à des modèles biomécaniques. A.T. Still, fondateur de l’ostéopathie dont nous célébrons les 150 ans, en avait d’ailleurs fait un fondement de son approche thérapeutique, se comparant à un ingénieur envisageant le corps humain comme un ensemble mécanique à rééquilibrer. Il est finalement apparu que la biomécanique opérant à l’échelle macroscopique des tissus, n’était pas suffisante pour appréhender les interactions systémiques complexes auquel la main du thérapeute se confronte lors de son action sur le corps de ses patients. Il fallait à la fois tenir compte de l’influence omniprésente et multiple des fonctions neurologiques qui câble l’ensemble de l’organisme et des régulations endocriniennes qui agissent aussi bien localement qu’à distance et rythme la vie des organismes complexes. Puis, au décours du XXIème siècle, avec les progrès technologiques permis par le numérique, la mécanobiologie a permis de révéler l’expression des forces mécaniques au sein du vivant. En révélant leur influence aux niveaux micro et nanoscopiques 1, 2, 3, 4, 5, 6, 7, 8, 9 aussi bien extra et intra-cellulaire jusqu’au noyau cellulaire 10, 11, 12, 13, sanctuaire génétique, elle en a dévoilé l’omniprésence en biologie.

En à peine trente ans, la mécanobiologie a fait la démonstration de la mécanosensibilité 14, 15, 16 comme fonction régulatrice essentielle à la vie cellulaire. Dans un premier temps, elle nous a montré l’implication de la membrane cellulaire et de protéines spécifiques transmembranaires comme les intégrines se liant aux éléments matriciels (collagènes, élastine, protéoglycanes), les cadhérines en interaction avec les membranes basales, ou les diverses protéines des jonctions serrées assurant la liaison intercellulaire, essentielles notamment aux structures épithéliales. Très vite, il est apparu que ces protéines membranaires ou transmembranaires, étaient en continuité avec l’espace intra-cellulaire et son cytosquelette composé de structures filamentaires fugaces comme les microtubules ou plus durables comme les filaments intermédiaires et les micro-filaments d’actine et de myosine. Ce réseau mécanique construit un ensemble dynamique, véritable continuum intracellulaire, communiquant mécaniquement avec l’ensemble des organites y compris le noyau dont les gènes expriment directement les contraintes mécaniques lorsqu’il est tracté ou comprimé. En réponse, elles expriment certains éléments impliqués dans les mécanismes de construction ou de résorption matricielle afin d’adapter le micro-environnement qu’elles habitent.

Cette mécanosensibilité des cellules désormais bien acceptée, il restait à en découvrir les interactions avec le métabolisme, tout particulièrement énergétique dont les mitochondries 17 occupent un rôle central, essentiel à leur adaptabilité au stress environnemental. Un premier article publié en 2022 par l’équipe du Pr Sirio Dupont 18 étudié ces propriétés mitochondriales en réponse à une modification de l’environnement matriciel. Ces chercheurs ont observé une augmentation de l’absorption de la cystine, induisant une activation du métabolisme du glutathion dans les cellules disposées sur une MEC molle. De même les cellules dont les propriétés contractiles ont été réduites ou inhibées, simulant ainsi les conditions d’une MEC molle, ont réagi de la même manière. En présence de conditions de contractilité réduite, ils ont également observé une augmentation générale de la production d’espèces réactives de l’oxygène (ERO). Ainsi l’augmentation du métabolisme du glutathion sur un support matriciel mou se trouve bien associé à une augmentation des ERO. Une analyse plus fine a montré par ailleurs que si l’on observe une augmentation des ERO aussi bien cytoplasmiques que mitochondriales, l’origine de la réponse cellulaire à une MEC molle est mitochondriale. Ils ont démontré également que la capacité respiratoire basale n’était pas altérée et que la phosphorylation oxydative n’était pas significativement atteinte mais, qu’en revanche, lorsque la respiration est accrue et que l’activité de NRF2 augmente, leur capacité respiratoire maximale se trouve diminuée. Enfin, les réseaux mitochondriaux se trouvaient altérés en cas de MEC molle avec une activation des processus de fission mitochondriale, une perturbation de la régulation des ERO et de NRF2, induisant une réponse anti-oxydante. L’origine de la réponse mitochondriale à une MEC molle serait liée à la formation de microfilaments d’actine du cytosquelette autour du réseau mitochondrial. En effet, des cellules en contact avec une MEC molle présentent une contractilité réduite du cytosquelette d’actomyosine sous-membranaire, mais la formation d’actine-F péri-mitochondriale, sous l’impulsion des protéines Spire1C et Arp2/3, ce qui favorise la fission des mitochondries. L’ensemble de ces données montraient une interaction entre les caractéristiques mécaniques matricielles et leurs conséquences métaboliques avec une cascade informationnelle agissant sur les structures cytosquelettiques, les réseaux mitochondriaux et le métabolisme énergétique cellulaire, notamment en cas de mobilisation accrue en réponse à un stress métabolique.

Les protéines motrices associées aux microtubules, la dynéine (en violet) et la kinésine (en rouge), forment le complexe moteur/adapteur avec les protéines Milton/TRAK1/2 (en jaune) et Miro (en orange), ces dernières étant insérées dans la membrane externe mitochondriale. La dynéine assure le transport vers le pôle − des microtubules, la kinésine vers le pôle +.
Les mitochondries peuvent également se déplacer le long des microfilaments d’actine grâce à différentes formes de myosine.
L’ensemble de ces résultats expérimentaux, qui ne portent que sur des cellules tumorales, ne répond certes pas encore aux multiples interactions métaboliques et hormonales auxquelles sont soumises l’ensemble des cellules de l’organisme. Cependant, ils commencent à redessiner un paysage biologique beaucoup plus complexe que celui exploré jusqu’à présent par la biochimie. Et surtout, ils amènent, en intégrant les forces mécaniques, à ne plus réduire les cellules à de simples colonies agglutinées dans une mise en culture expérimentale mais à toujours les envisager dans leur micro et macro-environnement matriciel. Si cela n’invalide pas totalement 150 ans de culture biologique basée quasi-exclusivement sur les forces électromagnétiques biochimiques, les auteurs nous invitent à réinterpréter systématiquement les conclusions des études en fonction du cadre dans lequel elles sont menées. De ce fait, il semble que la définition d’un nouveau système de communication cellulaire fondé sur la réception et la réponse aux forces mécaniques constitue d’ores et déjà une nécessité afin que les chercheurs et les enseignants l’intègrent à leur cadre conceptuel. Déjà un certain nombre de chercheurs ont assimilé une telle approche dans leurs problématiques de recherche que ce soit sur des aspects fondamentaux 19, 20,21 aussi bien, comme les auteurs de cet article 18 que pour élargir l’horizon des prises en charge corporelles en rééducation 22, en oncologie 23, dans la compréhension des processus cicatriciels 24, 25, 26, 27 ou en médecine spatiale 28. Enfin, nous suggérons que ce nouveau cadre conceptuel et expérimental mécanobiologique microscopique, complété par les avancées anatomiques macroscopiques du système fascial, doit inciter les thérapeutes manuels à se confronter à ces nouvelles connaissances pour réinterpréter les modèles actuellement en vigueur dans l’enseignement de leurs pratiques.
Bibliographie
- Sapir L, Tzlil S. Talking over the extracellular matrix: How do cells communicate mechanically? Semin Cell Dev Biol [En ligne]. 2017;71:99–105. Disponible: http://dx.doi.org/10.1016/j.semcdb.2017.06.010
- Moujaber O, Stochaj U. The cytoskeleton as regulator of cell signaling pathways. Trends Biochem Sci [En ligne]. 2020;45(2):96–107. Disponible: http://dx.doi.org/10.1016/j.tibs.2019.11.003
- Liebman C, McColloch A, Rabiei M, Bowling A, Cho M. Mechanics of the cell: Interaction mechanisms and mechanobiological models. Dans: Membrane Biomechanics. Elsevier; 2020. p. 143–84.
- Benn MC, Pot SA, Moeller J, Yamashita T, Fonta CM, Orend G, et al. How the mechanobiology orchestrates the iterative and reciprocal ECM-cell cross-talk that drives microtissue growth. Sci Adv [En ligne]. 2023;9(13):eadd9275. Disponible: http://dx.doi.org/10.1126/sciadv.add9275
- Alisafaei F, Mandal K, Swoger M, Yang H, Guo M, Janmey PA, et al. Vimentin intermediate filaments can enhance or abate active cellular forces in a microenvironmental stiffness-dependent manner [En ligne]. bioRxiv. 2022. Disponible: http://dx.doi.org/10.1101/2022.04.02.486829
- Wagh K, Ishikawa M, Garcia DA, Stavreva DA, Upadhyaya A, Hager GL. Mechanical regulation of transcription: Recent advances. Trends Cell Biol [En ligne]. 2021;31(6):457–72. Disponible: http://dx.doi.org/10.1016/j.tcb.2021.02.008
- Pocaterra A, Romani P, Dupont S. YAP/TAZ functions and their regulation at a glance. J Cell Sci [En ligne]. 2020;133(2):jcs230425. Disponible: http://dx.doi.org/10.1242/jcs.230425
- Romani P, Valcarcel-Jimenez L, Frezza C, Dupont S. Crosstalk between mechanotransduction and metabolism. Nat Rev Mol Cell Biol [En ligne]. 2021;22(1):22–38. Disponible: http://dx.doi.org/10.1038/s41580-020-00306-w
- Dupont S, Wickström SA. Mechanical regulation of chromatin and transcription. Nat Rev Genet [En ligne]. 2022;23(10):624–43. Disponible: http://dx.doi.org/10.1038/s41576-022-00493-6
- Alisafaei F, Jokhun DS, Shivashankar GV, Shenoy VB. Regulation of nuclear architecture, mechanics, and nucleocytoplasmic shuttling of epigenetic factors by cell geometric constraints. Proc Natl Acad Sci U S A [En ligne]. 2019;116(27):13200–9. Disponible: http://dx.doi.org/10.1073/pnas.1902035116
- Lityagina O, Dobreva G. The LINC between mechanical forces and chromatin. Front Physiol [En ligne]. 2021;12:710809. Disponible: http://dx.doi.org/10.3389/fphys.2021.710809
- Echarri A. A multisensory network drives nuclear mechanoadaptation. Biomolecules [En ligne]. 2022;12(3):404. Disponible: http://dx.doi.org/10.3390/biom12030404
- Gonzalez-Novo R, Zamora Carreras H, De Lope-Planelles A, Lopez-Menendez H, Roda-Navarro P, Monroy F, et al. 3D environment modulates persistent lamin distribution and the biomechanical signature of the nucleus [En ligne]. bioRxiv. 2023. Disponible: http://dx.doi.org/10.1101/2023.04.10.536202
- Huang G, Xu F, Genin GM, Lu TJ. Mechanical microenvironments of living cells: a critical frontier in mechanobiology. Acta Mech Sin. 2019;35(2):265-9.
- Argentati C, Morena F, Tortorella I, Bazzucchi M, Porcel- lati S, Emiliani C, et al. Insight into mechanobiology: How stem cells feel mechanical forces and orchestrate biological functions. Int J Mol Sci. 2019;20(21):5337.
- Chiquet M, Gelman L, Lutz R, Maier S. From mechanotransduction to extracellular matrix gene expression in fibroblasts. Biochim Biophys Acta. 2009;1793(5):911-20.
- Planet-Vie [En ligne]. Lefévère-Laoide J. La dynamique des mitochondries; 26 May 2023 [cité le 16 Jul 2024]. Disponible: https://planet-vie.ens.fr/thematiques/cellules-et-molecules/physiologie-cellulaire/la-dynamique-des-mitochondries
- Romani P, Nirchio N, Arboit M, Barbieri V, Tosi A, Michielin F, et al. Mitochondrial fission links ECM mechanotransduction to metabolic redox homeostasis and metastatic chemotherapy resistance. Nat Cell Biol [En ligne]. 2022;24(2):168–80. Disponible: http://dx.doi.org/10.1038/s41556-022-00843-w
- Brunet T, Bouclet A, Ahmadi P, Mitrossilis D, Driquez B, Brunet AC, et al. Evolutionary conservation of early mesoderm specification by mechanotransduction in Bilateria. Nat Commun [En ligne]. 2013;4(1):2821. Disponible: http://dx.doi.org/10.1038/ncomms3821
- Mammoto T, Mammoto A, Ingber DE. Mechanobiology and developmental control. Annu Rev Cell Dev Biol [En ligne]. 2013;29(1):27–61. Disponible: http://dx.doi.org/10.1146/annurev-cellbio-101512-122340
- Cao R, Tian H, Tian Y, Fu X. A hierarchical mechanotransduction system: From macro to micro. Adv Sci (Weinh) [En ligne]. 2024;11(11). Disponible: http://dx.doi.org/10.1002/advs.202302327
- Thompson WR, Scott A, Loghmani MT, Ward SR, Warden SJ. Understanding mechanobiology: Physical therapists as a force in mechanotherapy and musculoskeletal regenerative rehabilitation. Phys Ther [En ligne]. 2016;96(4):560–9. Disponible: http://dx.doi.org/10.2522/ptj.20150224
- Langevin HM, Keely P, Mao J, Hodge LM, Schleip R, Deng G, et al. Connecting (T)issues: How research in fascia biology can impact integrative oncology. Cancer Res [En ligne]. 2016;76(21):6159–62. Disponible: http://dx.doi.org/10.1158/0008-5472.can-16-0753
- Ng JL, Kersh ME, Kilbreath S, Knothe Tate M. Establishing the basis for mechanobiology-based physical therapy protocols to potentiate cellular healing and tissue regeneration. Front Physiol [En ligne]. 2017;8:303. Disponible: http://dx.doi.org/10.3389/fphys.2017.00303
- D’Urso M, Kurniawan NA. Mechanical and physical regulation of fibroblast–myofibroblast transition: From cellular mechanoresponse to tissue pathology. Front Bioeng Biotechnol [En ligne]. 2020;8. Disponible: http://dx.doi.org/10.3389/fbioe.2020.609653
- Kuehlmann B, Bonham CA, Zucal I, Prantl L, Gurtner GC. Mechanotransduction in wound healing and fibrosis. J Clin Med [En ligne]. 2020;9(5):1423. Disponible: http://dx.doi.org/10.3390/jcm9051423
- Dintwa L, Hughes CE, Blain EJ. Importance of mechanical cues in regulating musculoskeletal circadian clock rhythmicity: Implications for articular cartilage. Physiol Rep [En ligne]. 2023;11(15). Disponible: http://dx.doi.org/10.14814/phy2.15780
- Cialdai F, Risaliti C, Monici M. Role of fibroblasts in wound healing and tissue remodeling on Earth and in space. Front Bioeng Biotechnol [En ligne]. 2022;10:958381. Disponible: http://dx.doi.org/10.3389/fbioe.2022.958381